Siroshini Thiagarajan1,
Fui-Lu Yong2,
Hemaroopini Subramaniam2,
Vivien Yi-Mian Jong3,
Chan-Kiang Lim2,
Yee-How Say1 ![]()
For correspondence:- Yee-How Say Email: sayyh@utar.edu.my
Received: 16 November 2016 Accepted: 17 February 2017 Published: 31 March 2017
Citation: Thiagarajan S, Yong F, Subramaniam H, Jong VY, Lim C, Say Y. Anti-obesity effect of phenylcoumarins from two Calophyllum spp in 3T3-L1 adipocytes. Trop J Pharm Res 2017; 16(3):563-572 doi: 10.4314/tjpr.v16i3.9
© 2017 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
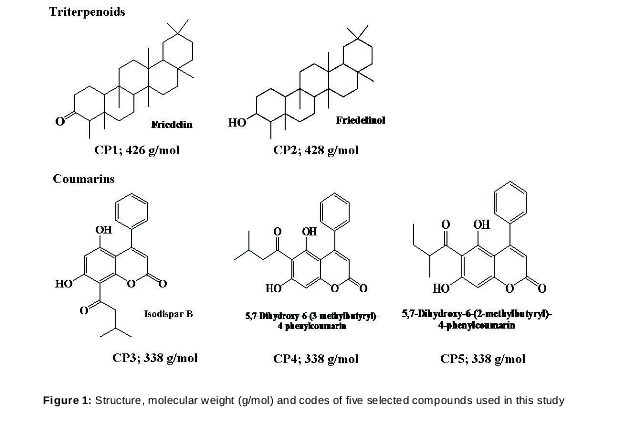
Purpose: To evaluate the anti-obesity effects of five compounds isolated from Calophyllum andersonnii and Calophyllum sclerophyllum, viz, friedelin (CP1), friedelinol (CP2), isodispar B (CP3), 5,7-dihydroxy-6-(3-methybutyryl)-4-phenylcoumarin (CP4) and 5,7-dihydroxy-6-(2-methybutyryl)-4-phenylcoumarin (CP5) in 3T3-L1 mouse pre-adipocytes.
Methods: Maximum non-toxic doses (MNTDs) of CP1 - CP5 were obtained by conducting 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Intracellular lipid droplet accumulation was determined by Oil Red O (ORO) staining. The effects of CP1 - 5 on the ex
Results: MNTDs for CP1-CP5 were 3.0, 1.4, 1.0, 29.0 and 25.0 µM, respectively. 3T3-L1 cells treated with CP1 - CP3 showed increased lipid accumulation (p < 0.05) and decreased glucose uptake (p < 0.05), compared with untreated cells; cells treated with CP4 and CP5 had opposite effects. Cells treated with CP4 and CP5 also showed downregulated Pparγ1, C/ebpα and aP2 ex
Conclusion: CP4 and CP5 isolated from Calophyllum sclerophyllum show promising anti-obesity properties, and could serve as candidate hits for further investigation at in vivo level to provide additional mechanistic evidence
Introduction
The genus Calophyllum, commonly known as “bintangor” or “penaga” in Malay, is a large group of tropical trees made up of around 200 different species [1]. Numerous Calophyllum species have been used in folk medicine. In Malaysia, the seed oil is used as a remedy for ulcer and rheumatism ailments. Moreover, Dweck and Meadows [2] reported the wound-healing property of Calophyllum seed oil after applying on scars. An infusion of the leaves is used to treat inflamed eyes.
Further pharmacological research on this genus has further revealed a variety of biological activities exhibited by these plants, such as anti-HIV, antibacterial, antimalarial, antioxidant, antitumor-promoting, and cytotoxic activities [3]. These biological activities have been attributed to phytochemicals such as xanthones, coumarins, chomanones (flavonoids, biflavonoids), terpenes and steroids [3].
Isolation of phytochemicals from Calophyllum andersonnii and Calophyllum sclerophyllum (native in the jungle of Landeh, Sarawak, Borneo Malaysia) yielded two classes, namely triterpenoids (friedelin, freidelinol) and coumarins (isodispar B, 5,7-Dihydroxy-6-(3-methybutyryl)-4-phenylcoumarin and 5,7-Dihydroxy-6-(2-methybutyryl)-4-phenylcoumarin)[4]. These phytochemicals could have potential adipogenic modulatory effects, as previous studies have shown that friedelin and freidelinol isolated from other plants have pro-adipogenic effects [5,6] while esculetin, a coumarin, has anti-adipogenic effect [7,8].
Therefore, the objective of this study was to investigate if the five triterpenoids and coumarins above would have the same pro-/anti-adipogenic effects on a differentiated mouse pre-adipocyte cell line, 3T3-L1. Adipogenic parameters assessed included intracelluar lipid droplet accumulation, glucose uptake, gene expression of adipogenic transcription factors and secretion of adipokines.
Methods
Plant, isolation and characterization of compounds
Triterpenoids - friedelin (compound 3) and friedelinol (compound 11) were isolated from Calophyllum andersonnii, while coumarins - isodispar B (compound 1), 5,7-Dihydroxy-6-(3-methybutyryl)-4-phenylcoumarin (compound 2) and 5,7-Dihydroxy-6-(2-methybutyryl)-4-phenylcoumarin] were isolated from Calophyllum sclerophyllum [4] The detailed steps involved in the isolation and characterization of the compounds used in this study are as described previously [4]. shows the structure, codes and molecular weights of the five compounds.
Cell culture
Mus musculus mouse fibroblast cell line 3T3-L1 (ATCC® CL-173™) was obtained from the American Type Culture Collection (Manassas, VA, U.S.A.). Methods to differentiate 3T3-L1 pre-adipocytes were achieved according to ATCC protocol. Seeded cells were maintained in pre-adipocyte expansion medium [90 % Dulbelcco’s Modified Eagle’s Medium (DMEM; Thermo Fischer Scientific PA, USA), 10 % Fetal Bovine Serum (FBS; Sigma-Aldrich, MO, USA)] by incubating them at 37 °C in a 5 % humidified CO2 incubator for 48 h until 100 % confluence was achieved (day 0). To induce differentiation, cells were incubated with differentiation medium [90 % DMEM, 10 % FBS, 0.5 mM 3-isobutyl-1-methylxanthine (Merck KGaA, Germany), 0.25 µM dexamethasone (Merck KGaA, Germany), 1 µg/mL insulin (Nacalai Tesque, Japan) and 2 µM rosiglitazone (Nacalai Tesque, Japan)] for 72 h (day 3). Cells were then maintained in adipocyte maintenance medium (90 % DMEM, 10 % FBS and 1 µg/mL insulin) for another 2 d (day 5), which was subsequently changed every 2 d. At day 10, the cells were fully differentiated after induction and subsequent bioassays were performed.
Determination of maximum non-toxic dose (MNTD)
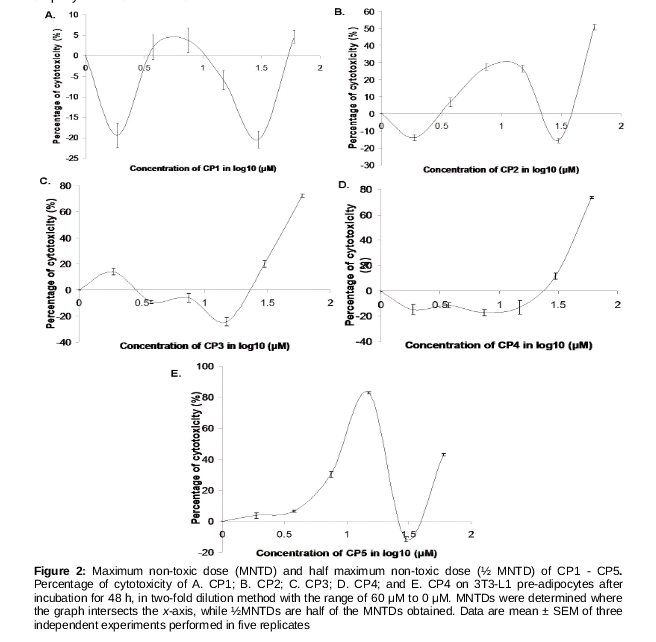
A total of 2500 3T3-L1 pre-adipocytes cells/well were plated in 96-well plates. At 70 % confluence, the cells were treated with the test compounds (0, 1.875, 3.75, 7.5, 15, 30 and 60 µM), dissolved in dimethyl sulfoxide (DMSO). Cell viability was assessed by 3-(4,5-dimethylthiazol-2-yl)-2.5-diphenyltetrazolium bromide (MTT) assay. The cells were incubated for 48 h at 37 ºC in humidified 5 % CO2. After 48 h, cells were rinsed twice with phosphate buffer saline (PBS). Twenty microliters of 5 mg/mL MTT (Bio Basic Canada Inc, Canada) stock solution were added to each well, further incubated at 37 ºC for 4 h, before the addition of 100 µL DMSO to solubilize the purple formazan crystals. After 1 h of incubation, the absorbance was measured at 570 nm using a microplate reader (M200 Tecan, Switzerland). A graph of percentage of toxicity against the log10 concentration of phytochemicals was plotted to determine the MNTD or ½ MNTD of the compounds. MNTD was determined at the first x-intercept where the toxicity is 0 %, while ½MNTD was half of the MNTD selected. These dosages were then added together in the differentiation medium at day 0 of differentiation as described in ‘Cell Culture’.
Determination of intracellular lipid droplet accumulation
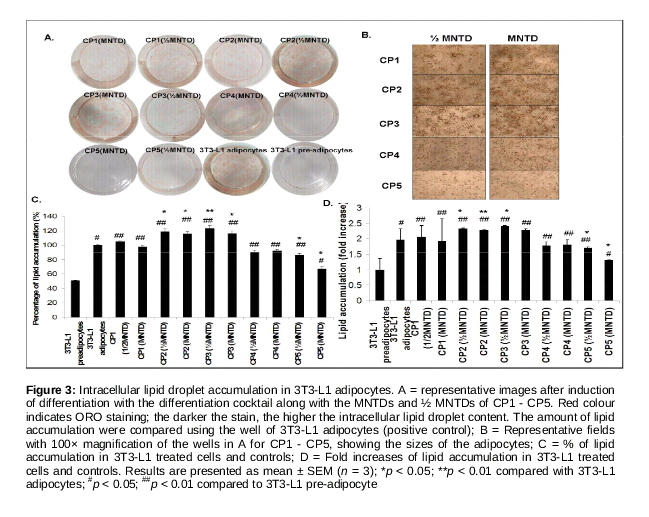
Eight days (day 10) after the induction of differentiation, the cells were rinsed three times with PBS. After fixation with paraformaldehyde (Sigma-Aldrich, MO, USA) for 1 h, the cells were rinsed with PBS and then stained with freshly diluted ORO solution [3 parts 0.5 % ORO (R&M Chemicals, UK) in isopropyl alcohol and 2 parts of water] for 30 min. The cells were then rinsed twice with water and visualized and photographed using an inverted phase contrast microscope (TS100, Nikon Eclipse, Japan). ORO stain was dissolved in isopropyl alcohol for quantitative analysis, and absorbance at 520 nm was measured using a microplate reader (M200, Tecan, Switzerland).
Glucose uptake assay
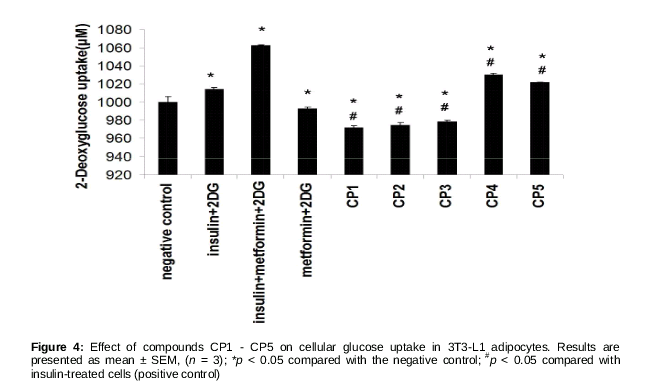
Glucose uptake was measured using glucose uptake assay kit (Abnova, Taiwan) according to manufacturer’s instructions. First, 3T3-L1 cells were plated in 96-well plate and differentiated, and at day 10 the differentiated cells were starved for 12 h without the addition of FBS to the basic medium. The cells were then treated with insulin (100 nM), metformin (1 mM; Sigma-Aldrich, MO, USA), MNTDs of CP1-CP5 and 2-deoxyglucose (2-DG) (negative control), for 1 h. Cellular 2-DG uptake was then measured by monitoring OD ratio increase at 570/610 using a microplate reader. The OD ratio for 2-DG (negative control) was normalised at 1000 µM.
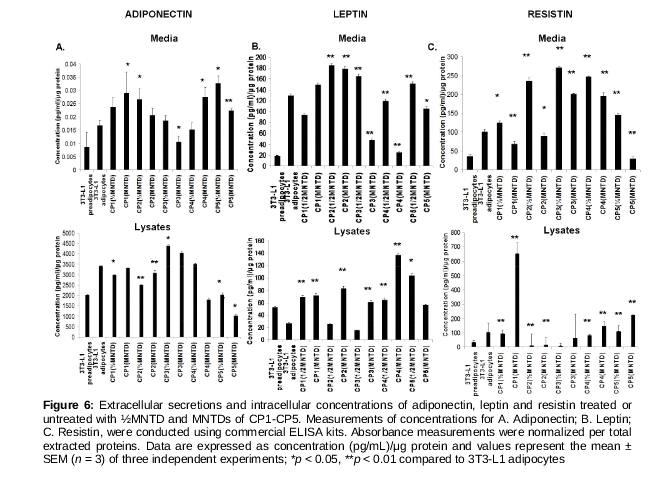
Measurement of adipokine concentration
After treatments, conditioned media were collected and cells were then rinsed with ice-cold PBS twice and lysed with 10 µL of lysis buffer [10 mM Tris/HCl, pH 7.8, 100 mM NaCl, 0.5 % (w/v) sodium deoxycholate, 0.05 % (v/v) nonidet-P40, 10 mM EDTA, 0.1 mM PMSF] added with 1 µL of protein inhibitor cocktail (Thermo Fischer Scientific PA, USA). Subsequently, concentrations of mouse leptin, resistin and adiponectin in the conditioned medium and cell lysates were determined using commercial ELISA kits (CUSABIO, China) according to manufacturer’s instructions. Protein concentrations of media and cell lysates were also measured by bicinchoninic acid protein assay kit (Thermo Fischer Scientific PA, USA) and adipokine concentrations were normalized per total extracted proteins.
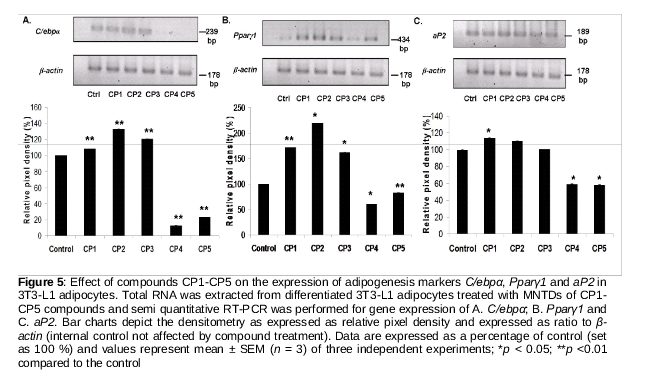
Determination of gene expression of adipogenesis markers
The gene expression of three common adipogenesis markers namely mouse nuclear receptor peroxisome proliferator-activated receptor gamma 1 (Pparγ1), CCAAT/enhancer binding protein-alpha (C/ebpα), and adipocyte fatty acid-binding protein (aP2) was assessed by semi-quantitative reverse transcription-polymerase chain reaction (RT-PCR). Total RNA was extracted from the 3T3-L1 adipocyte cells using EZ 10 Spin Column animal total RNA extraction kit (Bio Basic Canada Inc., Canada) according to manufacturer’s instructions. RNA purity and concentration were determined using a nanospectrophotometer (IMPLEM, Germany) at absorbance of 260 and 280 nm. The integrity of total RNA extracted was assessed by using 1 %(w/v) agarose gel electrophoresis and denaturing agent formamide. The primer sequences for RT-PCR were adopted from [8,9]. RT-PCR was performed using Realhelix™ RT-PCR Kit (Nanohelix, Korea) according to manufacturer’s instructions on Bio-Rad CFX-96T RT-PCR machine following this protocol: 1 cycle of cDNA synthesis at 55 °C for 50 min; 1 cycle of initial-denaturation at 95 °C for 15 min; 30 cycles of denaturation at 95 °C for 20 s, annealing at 53, 57, 57 and 62 °C for C/ebpα, Pparγ1, aP2, β-actin, respectively, for 40 s and extension at 72 °C for 40 s and a final cycle of post extension at 72 °C for 5 min. PCR products were resolved by 1.8 % agarose gel electrophoresis, stained with Gel Red® dye (Biotium, CA, USA) and visualized with BIO-RAD Gel Doc image analysis software (BIO-RAD Laboratories Inc., CA, USA). Densitometry was performed using NIH Image J software.
Statistical analysis
The results are expressed as mean ± standard error of the mean (SEM) of at least two independent experiments performed in at least triplicate, unless otherwise stated. Statistical analysis was performed using Student’s t-test for comparison between two means using IBM SPSS Statistics software version 16.0 (IBM, NY, USA). A p-value < 0.05 was considered as statistically significant.
Results
MNTD and ½ MNTD of the compounds
In the graph of cytotoxicity percentage in 3T3-L1 pre-adipocytes against log CP1-CP5 concentrations (), the percentage of cytotoxicity for CP1, CP2 and CP5 showed fluctuations across concentrations while the percentage of cytotoxicity for CP3 and CP4 showed an increasing trend across concentrations.. However, cell death was not evident at concentrations below 3.75 µM, for CP1 and CP2; and at concentrations below 30 µM for CP3 and CP4 (). MNTDs of CP1, CP2, CP3, CP4 and CP5 were 3.00, 1.44, 1.04, 29.0 and 25.0 µM, respectively, while their ½MNTDs were 1.50, 0.72, 0.52, 14.5 and 12.5 µM, respectively. Overall, cytotoxicity of CP3 was the highest, and triterpenoids CP1 and CP2 were up to 20-folds more cytotoxic than phenylcoumarins CP4 and CP5.
Phenylcoumarins inhibit intracellular lipid accumulation
As an evidence of adipogenesis, copious ORO dye stained material accumulated in differentiated 3T3-L1 adipocytes (positive control) while no staining was detected in 3T3-L1 preadipocytes (negative control, A). Compared to the negative control, cells treated with MNTDs and ½ MNTDs of CP1 - CP3 showed higher accumulation of dye-stained material; CP4 and CP5 showed opposite effects (A). A 100× magnification of the cells revealed larger adipocytes for CP1 - CP3 (indicating higher lipid droplet accumulation), while the opposite was observed for CP4 - CP5 (B).
As shown in C and D, there was almost 50 % less lipid accumulation in non-differentiated 3T3-L1 cells compared with differentiated cells. CP2 and CP3 at both doses significantly increased lipid accumulation compared with the positive control, while CP4 and CP5 at both doses decreased lipid accumulation of up to 0.7-fold, albeit with statistical significance for the latter compound. Taken together, this implies that CP4 and CP5 would indeed attenuate adipocyte differentiation.
Phenylcoumarins promote cellular glucose uptake in 3T3-L1 adipocytes
As shown in , treatment with insulin and with insulin + metformin (a type II diabetes mellitus drug) significantly increased cellular uptake of glucose in 3T3-L1 adipocytes. Treatment with MNTDs of CP1, CP2 and CP3 significantly decreased cellular glucose uptake compared with insulin-treated cells. On the other hand, CP4 and CP5 significantly stimulated cellular glucose uptake compared with insulin-treated cells, but still the amount was still lesser than the synergistic effect of insulin + metformin (). These indicate that phenylcoumarins CP4 and CP5 promote cellular glucose uptake in 3T3-L1 adipocytes, comparable with the effect of insulin.
Phenylcoumarins inhibit adipogenesis by reducing gene expression of C/ebpα, Pparγ1 and aP2
As shown in , cells treated with CP1, CP2 and CP3 presented significant increases in the expression of adipocyte-specific transcriptional factors when compared with the untreated cells.
Specifically, cells treated with CP2 had the highest expression of C/ebpα and Pparγ1. Adipocyte specific transcriptional factor, aP2 showed the highest expression when cells were treated with CP1, which was about 18 % higher compared to the control (). On the other hand, cells treated with CP4 and CP5 showed significant down-regulation of the expression of all three adipogenesis markers (), implying that CP4 and CP5 attenuate adipocyte differentiation.
Phenylcoumarins from increase adiponectin and leptin secretion and decrease resistin secretion
As shown in , differentiated 3T3-L1 adipocytes had increased secretions of adiponectin, leptin and resistin compared with undifferentiated pre-adipocytes. Cells treated with ½MNTD CP5 had the highest amount of adiponectin secretion in conditioned medium, consistent with the lowest amount of intracellular adiponectin (Figure. 6A). Leptin secretion in conditioned medium was the highest in cells treated with CP2 (½MNTD), while the same cells had among the lowest intracellular leptin concentration (B). Cells treated with CP4 had the highest intracellular leptin concentration. As for resistin, cells treated with CP3 (½MNTD) had the highest secretion (2.5 times higher) compared with differentiated adipocytes, while the lowest for cells treated with CP5 (MNTD) (C). For intracellular resistin concentration, cells treated with CP1 (MNTD) showed 6.5 times higher level compared with differentiated adipocytes (C).
Discussion
Previously, it was shown that friedelin, a triterpenoid isolated from Garcinia prainiana, upregulated adipocyte differentiation and glucose uptake in 3T3-L1 adipocytes [6], while esculetin (a coumarin derivative), either naturally isolated from Fraxinus rhynchophylla [7] or chemically-synthesized [8], has anti-adipogenic effect. In this study, we found that the anti-adipogenic and pro-glucose uptake activities of CP4 and CP5 are consistent with the reduced expression of adipogenesis markers Pparγ1, C/ebpα, aP2, increased secretion of adiponectin and reduced secretion of leptin and resistin.
It is widely reported that adipogenesis can be inhibited or promoted by various natural compounds [6-8]. Two out of three coumarins that showed anti-adipogenic activity are CP4 and CP5. Isodipar B (CP3), also a coumarin, however showed pro-adipogenic effect. This may be due to the structural differences within the coumarins, as reported in [7]. There were differences in the moiety of R-group added to coumarin, the backbone of the phytochemicals [6,10].
Methylbutyryl located on a different carbon might cause a difference in activity in adipocytes. In CP3, the R-group is located in C-8 of the coumarin backbone. These results therefore, suggest that methylbutyryl located in C-8 is important for exerting anti-adipogenic effect. For cells treated with CP1 and CP2, an induction in lipid accumulation was observed. Friedelin naturally isolated from Garcinia prainiana also exerted the same activity [6], albeit at a higher concentration of 10 μM (vs. 3 μM in our study).
Unlike friedelin from Garcinia prainiana which promoted cellular glucose uptake in 3T3-L1 adipocytes [6], our results showed the opposite effect. This discrepancy could be due to the higher dose used in our study. For the first time, we observed that treatment with CP4 and CP5 promoted 3T3-L1 adipocyte uptake of glucose, to the extent that it was even significantly higher than the positive control, insulin. Therefore, compounds CP4 and CP5 may imitate the action of insulin in stimulating cellular glucose uptake and it is speculated that these compounds may also reduce blood glucose level in vivo. Based on the above findings, it appears that our findings related to the effect of phenylcoumarins CP4 and CP5 on 3T3-L1 adipocytes were in agreement with two other reports [12,13] stating that compound that inhibits adipocyte differentiation could also improve glucose uptake.
In this study, we also measured the gene expression of three common adipogenesis markers C/ebpα, Pparγ1 and aP2 to see if their expression correlates with the adipocyte differention/lipid droplet accumulation as assessed by ORO staining. C/ebpα and Pparγ activate transcriptional adipogenesis activity by binding to their genomic promoter region interchangeably [14]. Cells treated with CP1, CP2 and CP3 showed pro-adipogenic activity, by increasing Pparγ1, C/ebpα and aP2 expression. Conversely, CP4- and CP5-treated cells showed a reduction in the expression of Pparγ1, C/ebpα and aP2. The findings are supported by previous studies which showed that the reduction in adipocyte differentiation and increase in cellular glucose uptake are also correlated with the downregulation of C/ebpα, Pparγ1 and aP2 expression [8,15].
Besides playing a role as an energy depot, the adipose tissue also secretes adipokines that are metabolic regulators. Three adipokines assessed in this study – adiponectin, leptin and resistin, modulate adipogenesis and are correlated with body mass index and total body fat [16]). Plasma levels of leptin and resistin has been shown to be proportional to body fat mass [17,18], whereas plasma adiponectin level is negatively correlated with the percentage of body fat [19]. Therefore, we hypothesized that CP4 and CP5 compounds which show anti-adipogenic/pro-glucose uptake effects would also show increased adiponectin and decreased leptin and resistin secretions in 3T3-L1 adipocytes. However, we found that CP1 - CP4 (MNTD) compounds produced mixed effects on the secretions of these adipokines, which do not correlate with their adipogenic/glucose uptake properties shown earlier. This phenomenon could be due to the unassessed complex signaling pathways that modulate the secretion of these adipokines and adipogenesis/glucose uptake independently. Nevertheless, CP5 (MNTD) consistently produced increased adiponectin and reduced leptin and resistin secretions, compared with untreated cells. Similarly, ginsenoside has been shown to reduce 3T3-L1 adipocyte lipid accumulation and increase adiponectin expression [20], while Lysimachia foenum-graecum treatment increased adiponectin level while decreasing leptin and resistin levels in the plasma of treated mice [21].
Conclusion
Friedelin and friedelinol isolated from Calophyllum andersonnii promote adipogenesis by increasing intracellular lipid droplet accumulation and expression of adipogenesis-related genes including Pparγ1, C/ebpα and aP2, and also by decreasing cellular glucose uptake in 3T3-L1 adipocytes. On the other hand, 5,7-dihydroxy-6-(3-methybutyryl)-4-phenylcoumarin and 5,7-dihydroxy-6-(2-methybutyryl)-4-phenyl-coumarin isolated from Calophyllum sclerophyllum show promising anti-obesity properties by attenuating adipogenesis (decreasing intracellular lipid droplet accumulation and expression of Pparγ1, C/ebpα and aP2), and also by increasing 3T3-L1 adipocyte glucose uptake. Last but not least, 5,7-dihydroxy-6-(2-methybutyryl)-4-phenylcoumarin increases adiponectin and reduces leptin and resistin secretions from 3T3-L1 treated cells.
Declarations
Acknowledgement
References
Archives


News Updates